Dairy & Veterinary Sciences - Juniper Publishers

Abstract
Heat stress (HS) has detrimental effects on lactating cattle especially when they are in a negative energy balance. Feed additives have been shown to mitigate the effects of HS by improving metabolic and immune function. The objective of this study was to evaluate the effect of feeding a dietary supplement (Land O’Lakes Inc., Arden Hills, MN) on the HS response in multi-parturient dairy cows in mid lactation. Two pens of cows at a commercial dairy were fed either control (CON) or additive (YB) at approximately 56.5 kg/pen per d for two weeks prior to arrival. Study cows (n=12) were balanced in days in milk (DIM), milk production, and parity (111.91±4.85 d, 33.67±0.96 kg/d, and 2.25±0.18). On arrival, the cows (6 YB and 6 CON) were randomly selected and housed in environmentally controlled chambers for 18 d and fed appropriate diet. Cows were subjected to 7 d of thermoneutral (TN) conditions, 7 d of HS, and 4 d of recovery (REC) under TN conditions. Dry matter intake (DMI), milk production from AM and PM milkings, and milk composition were measured daily. Rectal temperature (RT) and respiration rate (RR) were measured at peak temperature daily. Blood samples were collected once daily at 0900 h following catheterization on d 5 of TN to d 4 of REC. Serum samples were analyzed for glucose, insulin, blood urea nitrogen (BUN), β-hydroxybutyrate (BHB), and non-esterified fatty acids (NEFA). Results were analyzed using repeated measures in the PROC MIXED of SAS. HS increased RT (P<0.0001), RR (P<0.0001), BUN (P<0.0001), BHB (P=0.0009), insulin (P=0.036), cortisol (P<0.0001), neutrophil (P=0.0272), and water intake (P<0.0001). HS decreased lymphocyte (P<0.0001), DMI (P<0.0001), and 4% fat corrected milk (FCM, P=0.02). YB increased the feed efficiency ratio (P=0.03). YB had no effect on blood parameters. There was a treatment interaction with cows fed YB having higher feed efficiency (P=0.05) during peak thermal loads than CON. Results of this study suggest that HS exposure had performance and metabolic impacts in mid lactation cows. Supplementation with YB alleviated some of the performance effects associated with HS.
Keywords: β-hydroxybutyrate; Altering cortisol; Rumen acidosis; Ketosis; Lameness; Mastitis; Metritis
Abbreviations: HS: Heat stress; DIM: Days In Milk; TN: Thermoneutral; REC: Recovery; DMI: Dry Matter Intake; RT: Rectal Temperature; RR: Respiration Rate; BUN: Blood Urea Nitrogen; BHB: β-Hydroxybutyrate; NEFA: Non-Esterified Fatty Acids; THI: Temperature-Humidity Index
Introduction
HS is a costly factor to the dairy industry [1] as it changes the livestock production system due to increasing climate variability [2]. Physiological responses occur when the temperature-humidity index (THI) exceeds 68 or when the ambient temperature exceeds 32.2˚C [3]. Measurable factors that are negatively affected by heat stress are milk production, dry matter intake, growth, and health disorders [4]. It has been shown that milk yield is partially affected by HS through independent mechanisms of reduced nutrient intake [5].
Increasing the energy density of the diet from carbohydrates sources to fatty acid sources are common methods for feeding heat stressed animals but increase the chances of rumen acidosis per Kadzere et al. [6]. Using supplemental dietary additives have been shown to stabilize the rumen and increase post-rumen nutrient flow [7] in non-stressed cows; however, Shwartz et al. [8] reported no effect of a yeast culture on mitigating the effects of heat stress on lactating cows. The feed additive used in this study was a proprietary phytogenic and specialty yeast blend (Land O’Lakes Inc., Arden Hills, MN). Proven feed additives have shown increased immune function in lactating dairy cows [9] by altering cortisol responses in supplemented cows [10].
We hypothesized that supplementing with the YB would increase energy availability and thus reduce the negative effect of the heat load on production measures. The study objectives were to evaluate the effects of the dietary YB on body, production, and metabolic parameters in heat stressed lactating cows.
Materials and Methods
This study was conducted at the Agricultural Research Complex (ARC) at the University of Arizona (Tucson, AZ). The protocol was reviewed and approved by the University of Arizona Institutional Animal Care and Use Committee. It occurred in two phases: on-farm and on-site.
The on-farm phase consisted of feeding one 500 cow pen and monitoring a control pen of the same number of cows at a dairy in Stanfield, AZ. Treatment cows were given the feed additive in the TMR at 56.5 kg/pen and received it for 2 weeks. Previous research has shown that there may be a cell-mediated response in the rumen and that requires time to see a response during heat stress [10]. After the 2-week period, 6 treatment cows and 6 control cows were randomly selected and shipped to the ARC for the on-site phase with no health issues (rumen acidosis, ketosis, lameness, mastitis, metritis).
Upon arrival, twelve multiparous Holstein cows producing 33.67 ± 0.96 kg of milk/d, lactation (2.25 ± 0.18), and stage of lactation (111.92 ± 4.85 DIM) were weighed and randomly assigned to individual tie stalls in one of two environmental chambers. Both chambers housed 6 cows (3 control and 3 treatment). Chambers operated on the same environment throughout the study. There were 4 periods: acclimation (4 days), thermal neutral (TN, 7 days), heat stress (HS,7 days), and recovery period (4 days) (Figure 1). The acclimation period and TN period were set on the same environmental cycle. The TN environment was set at a minimum temperature of 21.16 ̊C, a maximum of 21.48 ̊C, and an average of 21.39 ̊C. The relative humidity (RH) had a minimum of 28.32 %, a maximum of 36.73 %, and an average of 32.30 %. The temperature-humidity index (THI) had a minimum of 65.54, a maximum of 66.16, and an average of 65.84 (Figure 2). The HS period had a minimum temperature of 25.40 oC, a maximum of 34.89 oC, and an average of 30.09 oC. The RH had a minimum of 23.40 %, a maximum of 36.48 %, and an average of 30.88 %. The THI had a minimum of 70.68, a maximum of 80.70, and an average of 75.46 (Figure 2). The Recovery period had a minimum temperature of 21.11 oC, a maximum of 21.43 oC, and an average of 21.38 oC. The RH had a minimum of 40.46 %, a maximum of 50.93 %, and an average of 45.93 %. The THI had a minimum of 66.39, a maximum of 66.99, and an average of 66.78 (Figure 2). The THI of 68 identifies as the threshold for HS in lactating dairy cows [11].


THI values were calculated using the average temperature and relative humidity obtained from the data logger. THI was defined by the formula [12]:
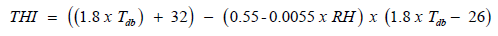
Where: db T = dry bulb temperature (̊oC), RH = relative humidity percentage.
The diet was an alfalfa based TMR that was balanced to be consistent with the green chop based TMR on the commercial dairy. Grab samples were collected once every 3 days when a new batch of feed was mixed. Alfalfa was added to the TMR pre-mix and was stored at 4 oC. Samples were analyzed by Dairy Nutrition Services INC (Chandler, AZ) by wet chemistry
Feeding and milking were done twice daily at 0530 and 1730 h. The CON cows were fed the base TMR. The YB cows were fed the TMR plus 113.4 g of YB mixed. Orts were removed and weighed in the morning prior to feeding. The feed efficiency was determined by using 4% fat corrected milk (FCM) per kilogram of DM consumed for each cow in each treatment group. Water consumption was recorded daily from the water meters after the AM feeding. Milk weights were recorded daily and a daily milk sample was taken from the AM milk. Milk samples were individually refrigerated with a preservative (Bronopol tablet, D&F CON Systems, San Ramon, CA) at 4 oC. Samples were analyzed for fat, lactose, protein, somatic cell count (SCC), and solids-not-fats (SNF) by Arizona DHIA (Tempe, AZ). Body weights were recorded twice during the study: once during HS and at the beginning of REC.
Physiological measurements such as respiration rate and rectal temperature were recorded for each cow once daily at 1400 h. Respiration rate was recorded as breaths per minute. It was calculated by counting the flank movements for 15s and multiplying by 4. Rectal temperature was measured using a Cooper TM99A thermometer (Cooper-Atkins, Middlefield, CT). Cows that exceeded 40.5oC were removed from the chamber and were cooled with water until a rectal temperature of 38.3oC. Vaginal temperatures were recorded with HOBO® U12 stainless steel temperature data loggers (Onset Computer Corp., Bourne, MA) in 5 minutes increments. Blank controlled internal drug releasing devices (Eazi-BreedTM CIDR®; Zoetis, Parsippany-Troy Hills, NJ) were used to hold the HOBO in the vagina. The cows were fit with this apparatus on d 1 for the duration of the study. CIDR placement was monitored and were refitted if they were expelled from the vagina.
Blood was collected via indwelling jugular catheters that were surgically inserted into cows on day 4 and 5. Blood was collected at 0900 h from day 5 to 18. Catheters were flushed before sampling and once at 1500 h with heparinized saline (100 USP/mL). The first 6 mL of fluid was discarded to eliminate heparin. Two 12 mL syringes with a 22-gauge needle were used to draw blood and were transferred to BD Vacutainer tubes (BD, Franklin Lakes, NJ): sodium heparin and blank. Samples were chilled for one hour at 4 oC. Serum and plasma were collected after centrifuging samples at 1,500 x g for 15 min at 4 oC and stored at -80 oC until analysis. Additional blood samples for white blood cell differentials were collected in BD Vacutainer tubes with EDTA. Differentials were run by Antech Diagnostics (Phoenix, AZ).
Serum β-hydroxybutyrate (BHB) was determined by a colorimetric kit (β-hydroxybutyrate (Ketone Body); Cayman Chemical, Ann Arbor,MI). Serum blood urea nitrogen (BUN) was determined using a colorimetric kit (DetectX Urea Nitrogen; Arbor Assays, Ann Arbor, MI). Serum glucose was measured using a colorimetric assay (Glucose Oxidase; Pointe Scientific Inc., Canton, MI). Serum non-esterified fatty acid (NEFA) was determined enzymatically through a commercial kit (Wako NEFA-HR(2); Wako Chemicals USA, Richmond, VA). Serum insulin was determined using an enzyme linked immunosorbent assay (ELISA) kit (Bovine Insulin, ELISA kit; ALPCO, Salem, NH).
Plasma cortisol was assessed using an enzyme immunoassay (EIA) kit (Cortisol Parameter Assay; R&D Systems Inc., Minneapolis, MN) with an intra- and interassay CV <5 %.
Statistical Analysis
Physiological and blood data were analyzed using a PROC MIXED model as a 2 x 2 replicated factorial design with the LSMEANS and PDIFF options within each environmental period as the REPEATED option with day within period as the repeated measure, and effect of the cow was nested within treatment, using a covariant structure. Rectal and vaginal temperatures were averaged per hour by day of trial. Environment (ENV), treatment (TRT), ENV x TRT, day(ENV), and treatment x day(ENV) effects were tested by means of the PDIFF option and were declared significant at P < 0.05. One YB cow was removed because of adaptation complications: severe hypophagia and constant standing.
Results and Discussion
Environmental conditions prior to arrival to ARC were not documented. All cows of the same treatment were housed in the same Saudi style barn cooled using Koral Kool systems and were kept below 72 THI. It isn’t known whether conditions were over 68 THI. The recorded environmental conditions for the TN, HS, and REC periods of the study are shown (Figure 2). As cows are exposed to conditions above 68 THI, RR exceeds 60 [13] and RT elevates above 38.5oC [3]. Due to these differences in parameters, changes in physiology and performance measures are prevalent between environmental groups Table 1.


Mean RR and RT in CON and YB fed cows are shown in Table 2. There was no difference in treatment for RR and RT. There is conflicting data on the effect of treatment on RT. Baumgard et al. [14] reported an increase in RT when cows were fed a feed additive during HS. Other studies reported a reduction in RT with supplementation during hyperthermia [9,15]. There was an effect of environment (P<0.0001) for both these variables as cows exposed to a THI above the threshold increases these parameters. HS cows increased RR (42 breaths/min) and RT (1.32 oC) compared to TN cows which indicates heat stress was achieved.
Feed intake, water intake, milk yield, and components are reported in Table 3. Dry matter intake (DMI) increased in both groups as cows adjusted to the facility during the TN period but experienced a slight decrease due to feed heating during 2 days of storage during TN period (Figure 3). DMI differed between TN and HS (P=0.0007). Thermal stress has been well documented in causing a decrease in DMI and how that drop in DMI may impact milk yield. Previous heat stress research has shown an altered endocrine profile that differs from nutrient restricted cows which affect precursors of milk components, such as glucose [14,16,5]. There was no effect of TRT and TRT x ENV for DMI. Schingoethe et al. [17] had similar findings in lactating Holsteins cows that were fed supplementation of a yeast culture had no effect on DMI during the summer. On the contrary, Wiedmeier et al. [18] suggested that during periods of stress, fungal supplemental products could be effective with increasing digestibility of structural carbohydrates. This is supported by Moallem et al. [19] using a live yeast supplementation which saw an increase in feed intake and efficiency during the hot season. There was no difference in water intake between treatment groups for each environmental phase (Figure 4). Water intake increased during HS from TN (P=0.0005) and decreased during REC from HS (P<0.0001). Murphy et al. [20] reported similar water intake values in lactating Holstein cows. Cows fed YB had a higher feed efficiency (1.34±0.08 vs. 1.15±0.08, P=0.05) compared to CON (Figure 5). There was also an effect of ENV (P<0.0001) with feed efficiency increasing during HS and decreasing during REC. Improvement in feed efficiency has been reported in hyperthermic conditions with supplementation of feed additives that contained live yeast and yeast culture [19,17]. YB increased feed efficiency in general even though there was no ENV effect. BW differed between TRT (P=0.0235) with YB cows having lower BW than CON. There was an effect of ENV (P=0.0103) with BW decreasing during periods of HS. The interaction between ENV x TRT was not significant for BW.

Milk and FCM yield differences weren’t detected between CON and YB groups (Figure 6 and Table 3). These results are similar to previous studies using feed additives containing yeast cultures [17,21]. Other studies have shown an increase in milk yield with yeast supplementation but it is correlated with greater DMI for ruminants [22,23]. Milk yield declined during HS (P=0.002). This decline in milk yield is only partially due to a decrease in DMI which has been shown by Rhoads et al. [16]. Differences between environments were found in FCM with TN being more elevated than HS (P=0.02). There was no significant difference in the following milk components: protein percentage, lactose percentage, and SCC. Previous work by Hall et al. [10] with HS has shown an increase in SCC during the REC but there was no significance observed in this study. The hypothesis is that during heat stress, mammary epithelial cells sluff post HS and contribute to the rise in SCC. Fat percentage had an effect of ENV (P=0.0129). Milk fat percentage had no difference between TN and HS which is surprising while there was fat depression observed during the REC. Rhoads et al. [16] reported that climate-controlled HS cows don’t have reduced milk fat percentages, unlike commercial dairies that frequently experience fat depression during the summer months due to supplementation with high concentrate diets [6]. Not much work has been done during the REC and further research needs to evaluate this in climate controlled cows.





Hormone and metabolite data are presented in Table 4. No differences were detected in serum glucose due to TRT, ENV or TRT x ENV (Figure 7). This is contrary to other findings [16,8]. Cowley et al. [24] had similar results and suggests that the glucose homeostatic regulatory system is able to compensate for the reduction of DMI. No significant differences in serum NEFA for TRT, ENV or TRT x ENV were detected. Serum BHB did not have any significant differences in TRT or TRT x ENV but an effect of ENV was observed (P=0.0009). NEFA and BHB are a significant source of energy for cows in a negative energy balance. BHB decreases during HS according to Dale and Brody [25] but a decrease during REC which has TN conditions is surprising. This suggests a potential shift in postabsorptive changes in lipid metabolism since ketones are a by-product of fatty acid oxidation and the cow now favors glucose to lower heat production.

There was a difference in ENV for BUN (P<0.0001) and there were no differences in effect for TRT or TRT x ENV. The increase of BUN during HS is similar to previous studies [8,5]. BUN originates from two sources: inefficient rumen ammonia incorporation or deamination of amino acids as glucose precursors. Erasmus et al. [7] found that yeast supplemented groups during HS had improved rumen nitrogen balance which supports the increase in urea-nitrogen in circulation. During periods of inadequate nutrition, cows will undergo proteolysis in skeletal muscle to support lactation [26]. BUN source could have derived from muscle catabolism; however, BUN isn’t a good marker and either 3-methyl-histidine or creatine should be used which increase in HS lactating cows [27,28].
No difference was detected in serum insulin between TRT groups and TRT x ENV. There was an effect of ENV (Figure 8) with insulin increasing during the HS period (P=0.036). Elevated insulin levels have been reported likewise during HS periods [29,5]. This helps explain why there is no NEFA response due to insulin’s antilipolytic properties [30]. Insulin also stimulates protein synthesis and serves as an antiproteolyic hormone [31] which supports the decrease in BUN.

Plasma cortisol had no effect of TRT and TRT x ENV. As expected, there was a significant impact of ENV on plasma cortisol (Figure 9; P<0.0001). Plasma cortisol has been noted to have an increase during acute but not chronic HS [32]. Interestingly, we found no response during the acute phase for acclimation to HS and an elevation during the chronic phase. Alvarez and Johnson [33] reported cortisol spikes that ranged from minutes to hours after an immediate insult of heat stress which would be on d 8. During acute heat stress, cows adjust cellular pathways to mitigate the effects of heat stress and maintain homeostasis. This is no longer needed once the acclimated phenotype and cows have reached long term heat acclimation [34]. This supports the decrease of cortisol during the acclimation period. One possibility is the collection of blood samples in the AM may have missed the spike in cortisol since cortisol levels peak in the afternoon. Another possibility is that although similar THI was used to previous studies conducted on HS, a difference in temperature and humidity could have caused this since there can be the same THI but different values for temperature and humidity

White blood cell differentials are shown in Table 5. Neutrophil count increased during HS (Figure 10) with an observable difference with an effect for ENV (P=0.02) but not for TRT and TRT x ENV. Lymphocytes decreased during HS and REC (Figure 11; P<0.0001) while having no other differences in TRT and TRT x ENV. Interestingly, there was no difference in the neutrophil to lymphocyte ratio. Total white blood cell count was more elevated during TN and decreased throughout the thermal periods respectively (ENV, P=0.009). The results agree with stress induces neutrophilia and lymphopenia [35,36-38,30].



In conclusion, supplementing mid-lactation Holstein cows with YB beginning 14 days before and during exposure to moderate HS showed an increase in some production responses measured by feed efficiency. Feeding YB; however, did not result in the mitigation of negative physiological responses associated with HS. The mechanism behind the increase in feed efficiency is unknown due to there being no glucose or DMI response to treatment and further research should focus on it.
Click here: https://juniperpublishers.com/index.php



Wow, that is quite informative. I like this article very much. The content was good. If any of the engineering students are looking for a projects for ece projects, I found this site and they are providing the best service to the engineering students regarding the projects ece projects
ReplyDelete